
Специфические рецепторы поперечнополосатых мышц[править | править код]
Помимо свободных нервных окончаний (ноцицепторов) и рецепторов соединительнотканных футляров в мышцах и области мышечно-сухожильных соединений имеется два типа важных рецепторов глубокой чувствительности:
- мышечные веретена;
- сухожильные органы Гольджи.
Эти рецепторы относятся к группе проприорецепторов, которые играют важную роль в выполнении движений и поддержании позы. Проприорецепторы передают информацию о позе тела в пространстве и расположении конечностей относительно туловища (чувство положения). Кроме этого, они регистрируют движения в суставах и их скорость (чувство движения) и могут оценивать приложенное мышечное усилие (чувство силы). Эта информация далее направляется вместе с импульсами от органов чувств и равновесия в высшие центры моторной деятельности (в том числе в мозжечок). Эти импульсы передают основную информацию для контроля движений и коррекции поддержания позы и целенаправленных движений.
Мышечные веретена[править | править код]
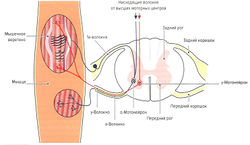
Рис. 1.6 а. Строение и функция мышечного веретена: кольцеспиральные нервные окончания 1а-волокон (Аа-волокна)
Мышечные веретена расположены по всей мышце. Их название обусловлено их веретенообразной формой. Количество мышечных веретен в мышце варьирует от 40 до 500 (Lindel, 2006). Как уже было отмечено, они лежат вдоль мышечных волокон (экстрафузальных волокон, вне веретена) и имеют 5-10 мм в длину и 0,2 мм в толщину (Kolster, 2006). Мышечные веретена состоят из соединительнотканной капсулы, поверхностной слой которой соответствует эндомизию, и 10-20 так называемых интрафузальных мышечных волокон (внутри веретена). Концы веретена прикрепляются к соединительнотканной оболочке мышцы или к сухожилию. Интрафузальные волокна только на концах имеют актиновые и миозиновые филаменты, позволяющие им сокращаться, а в центре волокно способно лишь к растяжению.
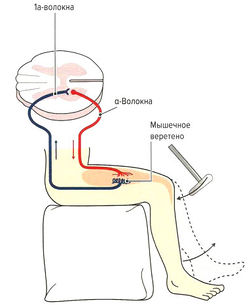
Рис. 1.6 б. Строение и функция мышечного веретена: моносинаптический проприоцептивный рефлекс
Несократительный растяжимый центр интрафузального волокна окружен толстым спиралевидным миелинизированным нервным окончанием типа 1а (Аа-волокна) (кольцеспиральное окончание) (рис. 1.6, а). Диаметр la-волокон составляет 10-20 мкм. Данные нервные волокна являются самыми быстрыми в организме человека — скорость распространения импульса составляет 60-120 м/с. Сократительные концы интрафузальных волокон иннервируются эфферентными у-волокнами (Ay-волокна, у-мотонейроны). Диаметр этих волокон составляет 4-8 мкм, а скорость передачи импульса — 20-50 м/с. Около 50 % всех мотонейронов передних рогов одного сегмента спинного мозга представлены у-мотонейронами (Cabri, 1999).
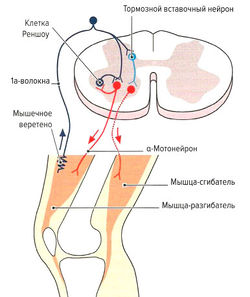
Рис. 1.6 в. Строение и функция мышечного веретена: реципрокное и возвратное ингибирование
Растяжение центральной части интрафузальных волокон приводит к раздражению кольцеспиральных нервных окончаний мышечного веретена (рецептор растяжения). На конце афферентных волокон возникает потенциал действия, который затем передается в ЦНС. Центральная часть веретена растягивается либо при растяжении всей мышцы, либо при напряжении сократительных концов интрафузальных волокон при активации у-мотонейронов. При одновременном действии этих двух факторов возможно как взаимное усиление, так и уменьшение их действия. Таким образом, мышечные веретена способны регистрировать как изменение длины мышцы, так и скорость ее изменения, а также косвенно ее изменять. Действуя через различные контуры регуляции, мышечные веретена выполняют несколько функций.
- Защита мышцы от перерастяжения за счет рефлекса растяжения (моносинаптический рефлекс). Функцией моносинаптического рефлекса растяжения является поддержание мышечной длины. Любое растяжение мышцы, например при ударе по сухожилию или мышце (рис. 1.6, б) или при изменении позы туловища, влияет на натяжение мышечных веретен. В результате резко повышается частота импульсации по 1а-афферентным нервным волокнам, идущим через задние спинномозговые корешки в серое вещество спинного мозга и через один синапс (моносинаптически) связываются с а-мотонейронами этой же мышцы (проприоцептивный рефлекс) (рис. 1.6, б). а-Мотонейроны по а-волокнам вызывают сокращение экстрафузальных волокон натянутой мышцы. Поскольку рецептор и эффектор находятся в одном и том же органе, скорость данного рефлекса очень высока (около 30 мс). Непроизвольные изменения длины мышцы могут сразу корректироваться на уровне спинного мозга без вовлечения высших моторных центров. Одновременно с этим через тормозной вставочный нейрон (ингибирующий la-нейрон) происходит ингибирование мотонейронов мышц-антагонистов (полисинаптический рефлекс) (реципрокное ингибирование антагонистов) (рис. 1.6, в). Благодаря этому растянутая мышца не преодолевает сопротивление антагонистов и предотвращается аналогичная реакция антагонистов при их растяжении (Weiss, 2000). После сокращения растянутой мышцы импульсация от la-волокон снижается. Дополнительно вследствие активации а-мотонейронов через коллатеральные связи и тормозные вставочные нейроны (клетки Реншоу) происходит обратное ингибирование самих а-мотонейронов (рис. 1.6, в). Эта отрицательная связь останавливает распространение сигнала и обеспечивает кратковременность сокращения мышцы, поэтому, несмотря на постоянные внешние раздражители (растяжение мышцы), возможно поддержание ее постоянной длины. Кроме этого, остановка рефлекторного ответа обеспечивается сухожильными органами Гольджи, которые активируются при мышечном сокращении и последующем растяжении сухожилия (аутогенное ингибирование) (см. рис. 1.7, б). В заключение необходимо отметить, что возбудимость мотонейронов контролируется высшими центрами головного мозга. При повреждении этих центров или отходящих от них нервных путей (например, при параплегии) развивается аномальное повышение рефлексов (см. рис. 1.6, а).
- Регуляция активного мышечного тонуса. Высшие моторные центры в головном мозге через так называемые экстрапирамидные пути активируют у-мотонейроны и через мышечные веретена регулируют длину мышц и, следовательно, активный мышечный тонус. Активация у-мотонейронов вызывает напряжение сократительных концов интрафузальных волокон, что растягивает центр мышечных веретен и приводит к активации la-волокон. Как уже было отмечено, импульс затем передается через задние рога в передние рога спинного мозга и возбуждает а-мотонейроны. а-Мотонейроны, в свою очередь, вызывают сокращение волокон скелетных
мышц, что опосредованно уменьшает натяжение центральной части веретен. Эта рефлекторная реакция продолжается до тех пор, пока 1а-волокна не будут регистрировать натяжение (у-петля). При нормальной позе (сидя, стоя или при ходьбе) в большинстве мышц, особенно в работающих против силы тяжести, регистрируется постоянная невысокая активность афферентных волокон мышечных веретен, что позволяет поддерживать определенный активный мышечный тонус. Мышцы, действующие против силы тяжести, например мышца, разгибающая позвоночник, в положении стоя находятся в постоянном небольшом натяжении, т. к. туловище под влиянием силы тяжести наклоняется несколько кпереди. За счет постоянной активации у-мотонейронов достигается дополнительное растяжение средней части мышечных веретен, что вызывает активацию высших моторных центров для поддержания мышечного тонуса. Благодаря постоянной фоновой афферентной импульсации во время обычных движений мышечные веретена могут воспринимать как укорочение, так и растяжение мышцы. При усилении или уменьшении активности у-мотонейронов изменяется чувствительность мышечных веретен и регулируется их активность при конкретных позе и движении. Таким образом, у-мотонейроны устанавливают в каждой конкретной ситуации оптимальную рабочую область мышечных веретен, и они могут регистрировать мышечную длину и скорость ее изменения. Это позволяет устанавливать и поддерживать постоянный активный мышечный тонус, а следовательно, поддерживать равновесие и положение суставов и определенную позу (чувство позы и опоры).
- Выполнение точных движений и регуляция тонкой моторики. Возможность регулировать чувствительность мышечных веретен, особенно вместе с произвольными движениями, играет большую роль. При произвольных движениях, например при разгибании в коленном суставе, происходит помимо активации а-мотонейронов и повышение активности у-мотонейронов. За счет а-у-коактивации устанавливается оптимальная длина мышечных волокон, чтобы поддерживать их чувствительность во время всего движения. При сокращении регистрируется растяжение мышцы и становятся возможными очень точные движения. Крохме того, в научной среде продолжаются дискуссии, являются ли точные движения конечностями исключительно результатом изменения у-активности. Супраспинальные нервные центры могут изменять у-активность в у-петле (изменение частоты импульсации по 1а-волокнам) и таким образом изменять активность связанных с ними а-мотонейронов (Weiss, 2005).
Запомните: Чем больше мышечных веретен в мышце, тем более точные и дифференцированные движения возможны при ее сокращении. Например, в мышцах кисти и глазного яблока находится большое количество проприорецепторов (Kolster, 2005).
Сухожильные органы Гольджи[править | править код]
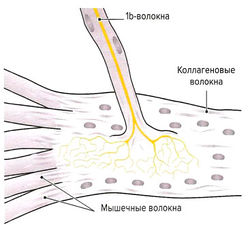
Рис. 1.7 а. Сухожильные органы Гольджи
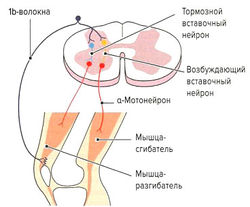
Рис. 1.7 б. Принцип аутогенного торможения
Помимо мышечных веретен, которые регулируют длину мышцы, в поперечнополосатой мышечной ткани находятся сухожильные органы Гольджи, регулирующие напряжение мышцы. Они располагаются вдоль экстрафузальных мышечных волокон в области мышечно-сухожильных соединений (рис. 1.7, а) и вплетаются в коллагеновые волокна сухожилия. Органы Гольджи состоят из множества разветвляющихся терминальных окончаний аксона lb-волокон (Аа-волокна), окруженных общей соединительнотканной капсулой. Эти рецепторы имеют в длину около 100 мкм и прикрепляются к 10-15 сухожильным волокнам (Cabri, 1999). При сокращении мышцы коллагеновые волокна натягиваются и сближаются. Расположенные между коллагеновыми волокнами нервные окончания сдавливаются и генерируют потенциал действия, который в виде нервного импульса передается по lb-волокнам в ЦНС. Адекватньш раздражителем для сухожильных органов Гольджи является изменение натяжения сухожилия, что происходит как при укорочении, так и при растяжении мышцы. Порог чувствительности этих рецепторов выше, чем у мышечных веретен. В отличие от моносинаптической дуги веретен, на уровне спинного мозга переключение импульса на а-мотонейрон идет через два или три синапса. Через тормозные вставочные нейроны (ди- или трисинаптически) происходит ингибирование а-мотонейронов заинтересованной мышцы аутогенное торможение). Одновременно с этим органы Гольджи через два синапса активируют через возбуждающие вставочные нейроны а-мотонейроны мышцы-антагониста (рис. 1.7, б). Оба этих механизма защищают мышцу от избыточного натяжения (защита от перегрузки).
Читайте также[править | править код]
- Поперечно-полосатые мышцы
- Кровоснабжение скелетных мышц
- Иннервация поперечно-полосатых мышц
- Cтроение и функции нейрона
- Синаптическая передача
- Нервно-мышечная передача
- Двигательная единица мыщцы
- Мышечно-сухожильное соединение
- Сухожилие
- Костно-сухожильное соединение
- Типы мышц
- Типы мышечных волокон
- Функции поперечно-полосатых скелетных мышц
- Гладкие мышцы
- Сердце
Рецепторы скелетной мышцы (мышечные веретена)
Можно выделить два основных вида специфических рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи). В этой статье рассмотрено строение и функции одного вида рецепторов скелетной мышцы — мышечных веретен. Описаны строение и функции интрафузальных мышечных волокон, входящих в состав мышечных веретен.
РЕЦЕПТОРЫ СКЕЛЕТНОЙ МЫШЦЫ (МЫШЕЧНЫЕ ВЕРЕТЕНА)
Познакомившись с строением нервов, можно обсудить вопрос о рецепторах скелетной мышцы. Можно выделить два основных вида рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи).
Мышечные веретена были описаны в середине XIX века немецким физиологом Вильгельмом Кюне (Wilhelm Kühne, 1863). Мышечное веретено представляет собой вытянутую структуру, расширенную посередине, что делает его похожим на веретено, которое в давние времена использовали, когда пряли. Английский физиолог, лауреат Нобелевской премии Чарльз Скотт Шеррингтон одним из первых указал, что мышечные веретена являются чувствительными рецепторами мышцы.
Мышечные веретена расположены внутри мышцы параллельно мышечным волокнам и прикрепляются к эндомизию, окружающему мышечные волокна или к сухожилию. Длина мышечного веретена достигает 10 мм. Ширина мышечного веретена в наиболее широкой его части — капсуле колеблется от 80 до 250 мкм (рис.1)
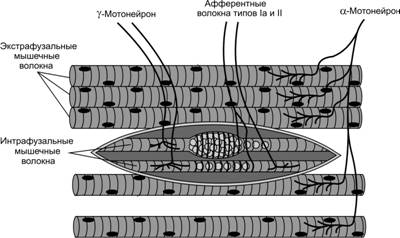
Рис.1. Строение мышечного веретена
Более подробно строение и функции мышц описаны в моих книгах
Количество мышечных веретен в мышце колеблется от нескольких десятков до нескольких сотен. Так, в двуглавой мышце плеча имеется 320 веретен, в большой грудной — 450, а в трехглавой мышце плеча — 520. Плотность веретен (то есть количество веретен приходящихся на 1 г массы мышцы) сильно варьирует. Она наибольшая в тыльных мышцах шеи и наименьшая в мышцах конечностей. Так, например в верхней косой мышце головы плотность веретен составляет 42,7 на 1 г массы мышцы, а в двуглавой мышце плеча — 2,0 на 1 г массы мышцы.
Установлено, что мышечные веретена возбуждаются при растягивании мышцы. Растяжение мышцы возникает, когда, например, занимающиеся выполняют упражнения на растяжение или упражнения в уступающем (эксцентрическом режиме). Также растяжение мышц возникает при локомоциях человека — ходьбе, беге, прыжках или езде на велосипеде.
Информация, поступающая от рецепторов мышечных веретен, сообщает нервной системе о том, какова в настоящий момент времени фактическая длина мышцы (ее еще называют текущей длиной — то есть длиной в текущий момент времени) и скорость растяжения мышцы.
Внутри мышечного веретена располагаются тонкие мышечные волокна, которые называются интрафузальными. Их диаметр в 2-3 раза меньше обычных мышечных волокон, называемых экстрафузальными. Интрафузальные мышечные волокна очень маленькие, однако они также могут сокращаться и расслабляться. Интрафузальных мышечных волокон в мышечном веретене немного: от 4 до 14. Следует отметить, что интрафузальные мышечные волокна в веретене имеют собственную иннервацию, которая позволяет регулировать чувствительность мышечного веретена. К интрафузальным мышечным волокнам подходят двигательные мотонейроны (γ-мотонейроны) и от них отходят чувствительные волокна, несущие информацию в ЦНС о длине и скорости растяжения мышцы. За открытия в этой области шведскому физиологу Рагнару Граниту была присуждена Нобелевская премия.
Благодаря информации, поступающей от мышечных веретен, предотвращается (правда не всегда) повреждение мышцы. Эта информация служит основой для функционирования рефлекса на растяжение.
Литература
- Гранит Р. Основы регуляции движений. — М.: Мир, 1973.- 278 с. Ил.
- Мак-Комас, А.Дж. Скелетные мышцы. — Киев: Олимпийская литература, 2001.- 407 с.
- Самсонова А.В. Моторные и сенсорные компоненты биомеханической структуры физических упражнений: автореф. дис… докт. пед. наук.- СПб, 1998.- 48 с.
- Самсонова, А.В. Биомеханика мышц [Текст]: учебно-методическое пособие /А.В. Самсонова Е.Н. Комиссарова /Под ред. А.В. Самсоновой /Санкт-Петербургский гос. Ун-т физической культуры им. П.Ф. Лесгафта.- СПб,: [б.н.], 2008.- 127 с.
- Самсонова, А.В. Гипертрофия скелетных мышц человека. — СПб: Кинетика, 2018.- 159 с.
С уважением, А.В.Самсонова
Похожие записи:
Повреждение и регенерация мышц. Эксперименты А.Н. Студицкого
Описаны эксперименты на животных, по повреждению и регенерации скелетных мышц. В этих экспериментах была показана высокая пластичность…
Томас ДеЛорме — ученый, разработавший метод прогрессивно возрастающего сопротивления
Описаны достижения врача-реабилитолога Томаса ДеЛорме, который разработал специальные силовые тренажеры для своих пациентов; использовал большие отягощения для борьбы…
Мышечные боли или почему болят мышцы после тренировок? (запаздывающие болезненные ощущения)
Описаны виды и причины болей в мышцах, возникающих через день после тренировки. Показано, что причиной мышечных болей…
Мышечные боли или почему болят мышцы во время и после тренировок? (острые болезненные ощущения)
Описаны виды и причины болей в мышцах после тренировки. Основное внимание уделено мышечным болям, возникающим во время…
От чего зависит сила мышц? (физиологические факторы)
Описаны физиологические факторы, определяющие силу скелетных мышц человека: частота импульсации ДЕ, количество активных ДЕ, синхронизация…
Классификация соматотипов детей по Штефко-Островскому
Описана классификация типов телосложения детей по Штефко-Островскому: астеноидного, торакального, мышечного и дигестивного. Кратко описан жизненный путь В.Г. Штефко.
Проприорецепторы. Мышечно-суставные рецепторы
 Необходимым условием нормальной мышечной деятельности является получение информации о положении тела в пространстве и о степени сокращения каждой из мышц. Эта информация поступает в центральную нервную систему от рецепторов вестибулярного аппарата, глаз, кожи, а также от проприорецепторов (мышечно-суставных рецепторов). К проприорецепторам относятся:
Необходимым условием нормальной мышечной деятельности является получение информации о положении тела в пространстве и о степени сокращения каждой из мышц. Эта информация поступает в центральную нервную систему от рецепторов вестибулярного аппарата, глаз, кожи, а также от проприорецепторов (мышечно-суставных рецепторов). К проприорецепторам относятся:
— мышечные веретена, находящиеся среди мышечных волокон,
— тельца Гольджи, расположенные в сухожилиях,
— пачиниевы тельца, находящиеся в фасциях, покрывающих мышцы, в сухожилиях, связках и периосте.
Все эти проприорецепторы относятся к группе механорецепторов. Мышечные веретена и тельца Гольджи возбуждаются при растяжении, а пачиниевы тельца — при давлении.
О важности афферентной иннервации мышц свидетельствует и тот факт, что 30-50% волокон в любом нерве, подходящем к мышце, являются афферентными, несущими импульсацию от проприорецепторов. К уже отмечалось, при выключении афферентной иннервации мышц, например при перерезке задних корешков спинного мозга, координированная мышечная деятельность нарушается, несмотря на сохранение двигательной иннервации мышц.
Мышечные веретена представляют собой высокодифференцированные рецепторные образования, снабженные афферентными и эфферентными нервными волокнами. Каждое веретено состоит из нескольких тонких так называемых интрафузальных поперечно-полосатых мышечных волокон.{_st-d-1}
Oдиночное волокно состоит из центральной части — ядерной сумки и двух способных к сокращению участков. В ядерной сумке pacположены рецепторы, представляющие сообой спиралевидные окончания толстых афферентных покрытых миелиновой оболочкой нервных волокон. Сокращающиеся участки интрафузального волокна иннервированы тонкими моторными, так называемыми гамма-эфферентными, волокнами, образующими мелкие концевые пластинки.
Один конец интрафузального волокна прикрепляется к обычному мышечному волокну, а другой — к сухожилию. Таким образом, интрафузальные волокна расположены параллельно массе мышечных волокон, среди которых они находятся. Если мышца растянута или расслаблена, то мышечные веретена тоже растянуты и в рецепторах ядерной сумки возникают импульсы, идущие в центральную нервную систему. Если же мышца сокращена, то натяжение мышечных веретен ослабевает и импульсация прекращается.
Второй вид проприорецепторов — тельца Гольджи — находится в сухожилиях. При мышечном сокращении тельца Гольджи растягиваются и в них возникают нервные импульсы, поступающие к нервным центрам. Сухожильные рецепторы менее возбудимы, чем мышечные веретена.
Таким образом, в двигательном аппарате имеются рецепторы — мышечные веретена,- возбуждающиеся при удлинении (расслаблении и растяжении), и рецепторы — тельца Гольджи,- возбуждающиеся при сокращении мышечных волокон.
Импульсы, поступающие в центральную нервную систему от мышечных веретен, облегчают возникновение рефлекторной реакции данной мышцы и тормозят сокращение мышцы-антагониста. Импульсы, приходящие от сухожильных рецепторов — телец Гольджи, вызывают противоположные рефлекторные реакции.
Импульсация из мышечных веретен, как показал Р. Гранит, может изменяться с помощью специального регуляторного механизма. Такой механизм образован сократительными элементами, расположенными по обе стороны от ядерной сумки интрафузального волокна. Их сокращение вызывает растяжение ядерной сумки и раздражение рецепторов веретена. Таким образом, эти рецепторы могут посылать в центральную нервную систему интенсивную импульсацию даже при небольшом сокращении или расслаблении поперечнополосатой мышцы.{_st-d-2}
Степень сокращения сократительных элементов веретена регулируется гамма-эфферентными нервными волокнами, которые являются отростками, гамма-мотонейронов спинного мозга. Импульсы, приходящие по гамма-эфферентным волокнам, вызывают сокращения интрафузальных мышечных волокон, что приводит к усилению потока афферентных импульсов от растягиваемых рецепторов ядерной сумки. Активность гамма-мотонейронов регулируется ретикулярной формацией среднего мозга.
Сократительные элементы мышечных веретен всегда находятся в состоянии некоторого тонуса, так как по гамма-эфферентам непрерывно поступают к ним импульсы из центральной нервной системы. Это влечет собой афферентную импульсацию от мышечных веретен, которая в свою ередь тонизирует мотонейроны спинного мозга, что является одной из причин рефлекторного тонуса скелетной мускулатуры. Отличительная особенность проприорецепторов — их малая способность к адаптации, благодаря чему центральная нервная система получает непрерывные сигналы о состоянии скелетной мускулатуры и может осуществлять непрерывную регуляцию двигательных актов.
Кинестетические сигналы, т. е. сигналы о движении отдельной части тела, играют чрезвычайно важную роль в развитии восприятий организма. Они являются главным контролем других органов чувств, например, зрения. Так, оценка зрением расстояния до какого-нибудь предмета вырабатывается при помощи мышечного чувства. Возбуждения, связанные с работой мышц, возникающие при хватании предмета руками или приближении к нему, сочетаются со зрительными ощущениями и служат важнейшим критерием для выработки суждения о том, насколько отдален предмет.
- Moustafine R. I., Bukhovets A. V., Sitenkov A. Y., Kemenova V. A., Rombaut P., Van den Mooter G. Eudragit® E PO as a complementary material for designing oral drug delivery systems with controlled release properties: comparative evaluation of new interpolyelectrolyte complexes with countercharged Eudragit® L 100 copolymers. Molecular Pharmaceutics. 2013; 10(7): 2630–2641. DOI: 10.1021/mp4000635.
- Киржанова Е. А., Хуторянский В. В., Балабушевич Н. Г., Харенко А. В., Демина Н. Б. Методы анализа мукоадгезии: от фундаментальных исследований к практическому применению в разработке лекарственных форм. Разработка и регистрация лекарственных средств. 2014; 3(8): 66–80. DOI: 10.33380/2305-2066-2019-8-4-27-31.
- Wise, «Review of the History of Medicine» (Л., 1967).
- https://sportwiki.to/%D0%A0%D0%B5%D1%86%D0%B5%D0%BF%D1%82%D0%BE%D1%80%D1%8B_%D0%BF%D0%BE%D0%BF%D0%B5%D1%80%D0%B5%D1%87%D0%BD%D0%BE-%D0%BF%D0%BE%D0%BB%D0%BE%D1%81%D0%B0%D1%82%D1%8B%D1%85_%D0%BC%D1%8B%D1%88%D1%86.
- https://allasamsonova.ru/receptory-skeletnoj-myshcy-myshechnye-veretena/.
- https://sport-51.ru/article/physiology/7464-proprioreceptory-myshechno-sustavnye-receptory.html.
- Bangun H., Aulia F., Arianto A., Nainggolan M. Preparation of mucoadhesive gastroretentive drug delivery system of alginate beads containing turmeric extract and anti-gastric ulcer activity. Asian Journal of Pharmaceutical and Clinical Research. 2019; 12(1):316–320. DOI: 10.22159/ajpcr.2019.v12i1.29715.
- Киржанова Е. А., Хуторянский В. В., Балабушевич Н. Г., Харенко А. В., Демина Н. Б. Методы анализа мукоадгезии: от фундаментальных исследований к практическому применению в разработке лекарственных форм. Разработка и регистрация лекарственных средств. 2014; 3(8): 66–80. DOI: 10.33380/2305-2066-2019-8-4-27-31.

")







