
Проприоцепция — это группа сигналов, посылаемых в центральную нервную систему специальными терминалами (проприоцепторами), расположенными в суставных капсулах, связках, сухожилиях и мышцах.
Проприоцепторы, среди которых выделяют мышечные рецепторы, или мышечные веретена (рецепторы растяжения), сухожильные рецепторы, или органы Гольджи (рецепторы мышечной силы), а также суставные рецепторы относятся к механорецепторам, посылающим в ЦНС информацию о положении, деформации и смещениях различных частей тела.
Функционирование этих рецепторов обеспечивает координацию всех подвижных органов и тканей животного и человека в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов животные теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
Если человек закроет глаза и попытается написать текст, то буквы будут написаны достаточно четко. Этим простым способом легко убедиться в умении человека пользоваться информацией, идущей от мышц и суставов.
Проприоцепторы составляют периферическую часть проприоцептивной сенсорной системы, или двигательного анализатора. Вместе с тем, несмотря на то, что миллионы людей ежедневно пользуются услугами этого анализатора, мы до сегодняшнего дня знаем сравнительно мало о его деятельности. Это особенно касается работы коркового отдела двигательного анализатора. Внутренние проприоцепторы находятся в мышцах, сухожилиях, сухожильных влагалищах, межкостных мембранах, фасциях, тканях суставов, надкостнице и т.д. Среди них имеются неспециализированные рецепторы, встречающиеся и в других частях тела (свободные нервные окончания, инкапсулированные рецепторы типа телец Руффини и Пачини), и специализированные — мышечные веретена и сухожильные органы (или рецепторы) Гольджи.
Двигательный, или кинестетический, анализатор (мышечная сенсорная система) обеспечивает формирование так называемого мышечного чувства при изменении напряжения мышц, суставных сумок, связок и сухожилий. Проблема мышечно-суставных ощущений имеет исключительное значение для физиологии и психологии. Специфические особенности человека появились благодаря более совершенной организации мышечной сенсорной системы по сравнению с животными.
И.М. Сеченов считал, что мышечное чувство является ближайшим регулятором движений и одним из орудий ориентации человека в пространстве и времени. Ощущения положения и перемещения тела в пространстве, ощущения во время трудовой деятельности и членораздельная речь лежат в основе формирования сознания человека и его представлений об окружающем мире.
Мышечное чувство обладает тремя качествами. Это, во-первых, ощущение положения конечностей, когда человек может определить положение своих конечностей и их частей относительно друг друга. Во-вторых, ощущение движения, когда, изменяя угол сгибания в суставе, человек осознаёт скорость и направление движения. Третьим качеством является ощущение усилия, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определённом положении при подъёме или перемещении груза.
Наряду с кожной, зрительной и вестибулярной сенсорными системами двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности.
Двигательная сенсорная система служит для анализа состояния двигательного аппарата — его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз.
Общий план организации. Двигательная сенсорная система состоит из следующих 3-х отделов:
- периферический отдел, представленный проприорецепторами, расположенными в мышцах, сухожилиях и суставных сумках;
- проводниковый и отдел, который начинается биполярными клетками (первыми нейронами), тела которых расположены вне ЦНС — в спинномозговых узлах. Один их отросток связан с рецепторами, другой входит в спинной мозги передает проприоцептивные импульсы ко вторым нейронам в продолговатый мозг(часть путей от проприорецепторов направляется в кору мозжечка), а далее к третьим нейронам — релейным ядрам таламуса (в промежуточный мозг);
- корковый отдел находится в передней центральной извилине коры больших полушарий.
Функции проприорецепторов
К проприорецепторам относятся мышечные веретена, сухожильные органы (или органы Гольджи) и суставные рецепторы (рецепторы суставной капсулы и суставных связок). Все эти рецепторы представляют собой механорецепторы, специфическим раздражителем которых является их растяжение.
Мышечные веретена человека, представляют собой продолговатые образования длиной несколько миллиметров, шириной десятые доли миллиметра, которые расположены в толще мышцы. В разных скелетных мышцах число веретен на1 г ткани варьирует от нескольких единиц до сотни.
Таким образом, мышечные веретена как датчики состояния силы мышцы и скорости ее растяжения реагируют на два воздействия: периферическое — изменение длины мышцы, и центральное — изменение уровня активации гамма-мотонейронов. Поэтому реакции веретен в условиях естественной деятельности мышц довольно сложны. При растяжении пассивной мышцы наблюдается активация рецепторов веретен; она вызывает миотатичес-кий рефлекс, или рефлекс на растяжение. При активном сокращении мышцы уменьшение ее длины оказывает на рецепторы веретена дезактивирующее действие, а возбуждение гамма-мотонейронов, сопутствующее возбуждению альфа-мотонейронов, приводит к реактивации рецепторов. Вследствие этого импульсация от рецепторов веретен во время движения зависит от длины мышцы, скорости ее укорочения и силы сокращения.
Сухожильные органы (рецепторы) Гольджи человека располагаются в зоне соединения мышечных волокон с сухожилием, последовательно по отношению к мышечным волокнам.
Сухожильные органы представляют собой структуру вытянутой веретенообразной или цилиндрической формы, длина которой у человека может достигать1 мм. Этот первичночувствующий рецептор. В условиях покоя, т.е. когда мышца не сокращена, от сухожильного органа идет фоновая импульсация. В условиях мышечного сокращения частота импульсации возрастает прямо пропорционально величине мышечного сокращения, что позволяет рассматривать сухожильный орган как источник информации о силе, развиваемой мышцей. В тоже время сухожильный орган слабо реагирует на растяжение мышцы.
В результате последовательного крепления сухожильных органов к мышечным волокнам (а в ряде случаев — к мышечным веретенам), растяжение сухожильных механорецепторов происходит при напряжении мышц. Таким образом, в отличие от мышечных веретен, сухожильные рецепторы информируют нервные центры о степени напряжения мыши, и скорости его развития.
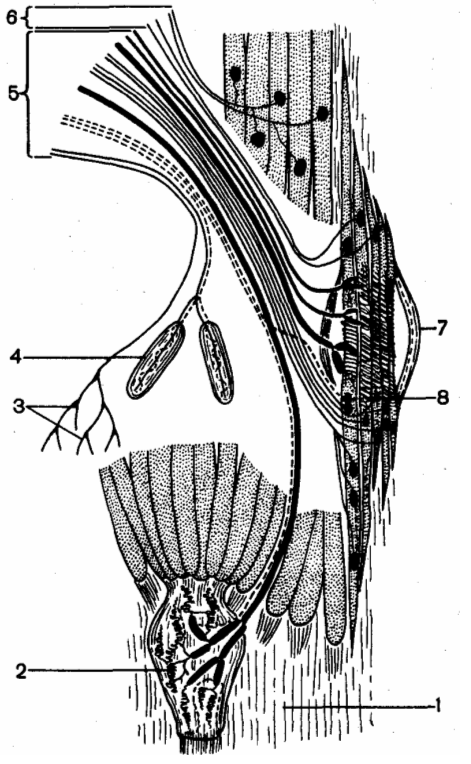
Эфферентная иннервация скелетной мышцы: 1 — сухожилие мышцы; 2 — сухожильные рецепторы Гольджи; 3 — свободные окончания; 4 — тельца Фатера-Пачини; 5 — афферентные волокна; 6 — эфферентные волокна; 7 — мышечное веретено; 8 — окончания афферентных волокон
Суставные рецепторы реагируют на положение сустава и на изменения суставного угла, участвуя, таким образом, в системе обратных связей от двигательного аппарата и в управлении им. Суставные рецепторы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецепторы посылают информацию о величине суставного угла, т. е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецепторы возбуждаются только в момент движения в суставе, т. е. посылают информацию о скорости движения. Частота их импульсации возрастаете увеличением скорости изменения суставного угла.
Проводниковый и корковый отделы проприоцеп-тивного анализатора млекопитающих и человека. Информация от мышечных, сухожильных и суставных рецепторов поступает через аксоны первых афферентных нейронов, находящихся в спинномозговых ганглиях, в спинной мозг, где частично переключается на альфа-мотонейроны или на вставочные нейроны (например, на клетки Реншоу), а частично направляется по восходящим путям в высшие отделы головного мозга. В частности, по путям Флексига и Говерса проприоцептивная импульсация доставляется к мозжечку, а по пучкам Голля и Бурдаха, проходящим в дорсальных канатиках спинного мозга, она доходит до нейронов одноименных ядер, расположенных в продолговатом мозге.
Аксоны таламических нейронов (нейронов третьего порядка) оканчиваются в коре больших полушарий, главным образом, в соматосенсорной коре (постцентральная извилина) и в области сильвие-вой борозды (соответственно, участки S-1 и S-2), а также частично в двигательной (префронтальной) области коры. Эта информация используется двигательными системами мозга достаточно широко, в том числе для принятия решения о замысле движения, а также для его реализации. Кроме того, у человека на основе про-приоцептивной информации формируются представления о состоянии мышц и суставов, а также, в целом, о положении тела в пространстве.
Сигналы, идущие от рецепторов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецепторов кожи, называют кинестетическими, т. е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецепторов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприорецепторов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз.
Таким, образом, проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза.
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов.
Вестибулярный анализатор — анализатор, обеспечивающий анализ информации о положении и перемещениях тела в пространстве.
https://www.braintools.ru/article/9907
Рецепторы скелетной мышцы (мышечные веретена)
Можно выделить два основных вида специфических рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи). В этой статье рассмотрено строение и функции одного вида рецепторов скелетной мышцы — мышечных веретен. Описаны строение и функции интрафузальных мышечных волокон, входящих в состав мышечных веретен.
РЕЦЕПТОРЫ СКЕЛЕТНОЙ МЫШЦЫ (МЫШЕЧНЫЕ ВЕРЕТЕНА)
Познакомившись с строением нервов, можно обсудить вопрос о рецепторах скелетной мышцы. Можно выделить два основных вида рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи).
Мышечные веретена были описаны в середине XIX века немецким физиологом Вильгельмом Кюне (Wilhelm Kühne, 1863). Мышечное веретено представляет собой вытянутую структуру, расширенную посередине, что делает его похожим на веретено, которое в давние времена использовали, когда пряли. Английский физиолог, лауреат Нобелевской премии Чарльз Скотт Шеррингтон одним из первых указал, что мышечные веретена являются чувствительными рецепторами мышцы.
Мышечные веретена расположены внутри мышцы параллельно мышечным волокнам и прикрепляются к эндомизию, окружающему мышечные волокна или к сухожилию. Длина мышечного веретена достигает 10 мм. Ширина мышечного веретена в наиболее широкой его части — капсуле колеблется от 80 до 250 мкм (рис.1)
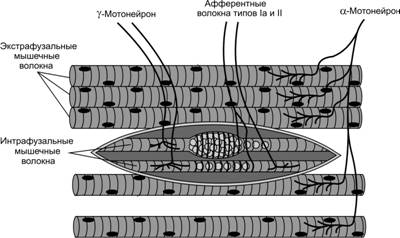
Рис.1. Строение мышечного веретена
Более подробно строение и функции мышц описаны в моих книгах
Количество мышечных веретен в мышце колеблется от нескольких десятков до нескольких сотен. Так, в двуглавой мышце плеча имеется 320 веретен, в большой грудной — 450, а в трехглавой мышце плеча — 520. Плотность веретен (то есть количество веретен приходящихся на 1 г массы мышцы) сильно варьирует. Она наибольшая в тыльных мышцах шеи и наименьшая в мышцах конечностей. Так, например в верхней косой мышце головы плотность веретен составляет 42,7 на 1 г массы мышцы, а в двуглавой мышце плеча — 2,0 на 1 г массы мышцы.
Установлено, что мышечные веретена возбуждаются при растягивании мышцы. Растяжение мышцы возникает, когда, например, занимающиеся выполняют упражнения на растяжение или упражнения в уступающем (эксцентрическом режиме). Также растяжение мышц возникает при локомоциях человека — ходьбе, беге, прыжках или езде на велосипеде.
Информация, поступающая от рецепторов мышечных веретен, сообщает нервной системе о том, какова в настоящий момент времени фактическая длина мышцы (ее еще называют текущей длиной — то есть длиной в текущий момент времени) и скорость растяжения мышцы.
Внутри мышечного веретена располагаются тонкие мышечные волокна, которые называются интрафузальными. Их диаметр в 2-3 раза меньше обычных мышечных волокон, называемых экстрафузальными. Интрафузальные мышечные волокна очень маленькие, однако они также могут сокращаться и расслабляться. Интрафузальных мышечных волокон в мышечном веретене немного: от 4 до 14. Следует отметить, что интрафузальные мышечные волокна в веретене имеют собственную иннервацию, которая позволяет регулировать чувствительность мышечного веретена. К интрафузальным мышечным волокнам подходят двигательные мотонейроны (γ-мотонейроны) и от них отходят чувствительные волокна, несущие информацию в ЦНС о длине и скорости растяжения мышцы. За открытия в этой области шведскому физиологу Рагнару Граниту была присуждена Нобелевская премия.
Благодаря информации, поступающей от мышечных веретен, предотвращается (правда не всегда) повреждение мышцы. Эта информация служит основой для функционирования рефлекса на растяжение.
Литература
- Гранит Р. Основы регуляции движений. — М.: Мир, 1973.- 278 с. Ил.
- Мак-Комас, А.Дж. Скелетные мышцы. — Киев: Олимпийская литература, 2001.- 407 с.
- Самсонова А.В. Моторные и сенсорные компоненты биомеханической структуры физических упражнений: автореф. дис… докт. пед. наук.- СПб, 1998.- 48 с.
- Самсонова, А.В. Биомеханика мышц [Текст]: учебно-методическое пособие /А.В. Самсонова Е.Н. Комиссарова /Под ред. А.В. Самсоновой /Санкт-Петербургский гос. Ун-т физической культуры им. П.Ф. Лесгафта.- СПб,: [б.н.], 2008.- 127 с.
- Самсонова, А.В. Гипертрофия скелетных мышц человека. — СПб: Кинетика, 2018.- 159 с.
С уважением, А.В.Самсонова
Похожие записи:
Томас ДеЛорме — ученый, разработавший метод прогрессивно возрастающего сопротивления
Описаны достижения врача-реабилитолога Томаса ДеЛорме, который разработал специальные силовые тренажеры для своих пациентов; использовал большие отягощения для борьбы…
Мышечные боли или почему болят мышцы после тренировок? (запаздывающие болезненные ощущения)
Описаны виды и причины болей в мышцах, возникающих через день после тренировки. Показано, что причиной мышечных болей…
Мышечные боли или почему болят мышцы во время и после тренировок? (острые болезненные ощущения)
Описаны виды и причины болей в мышцах после тренировки. Основное внимание уделено мышечным болям, возникающим во время…
От чего зависит сила мышц? (физиологические факторы)
Описаны физиологические факторы, определяющие силу скелетных мышц человека: частота импульсации ДЕ, количество активных ДЕ, синхронизация…
Классификация соматотипов детей по Штефко-Островскому
Описана классификация типов телосложения детей по Штефко-Островскому: астеноидного, торакального, мышечного и дигестивного. Кратко описан жизненный путь В.Г. Штефко.
Типы телосложения (соматотипы) по Хит-Картеру
Описаны типы телосложения (соматотипы) по классификации Б.Х. Хит и Д.Э.Л. Картера. Показаны преимущества разработанной классификации по сравнению…
ПРОПРИОЦЕПТОРЫ
Проприоцепторы (лат. proprius собственный, особенный + [ре]цепторы) — сенсорные приборы опорно-двигательного аппарата (мышц, сухожилий, фасций, суставов и др.). Иногда к Проприоцепторам относят также рецепторы вестибулярного аппарата и сердечнососудистой системы.
Первоначально термином «проприоцепторы», введенным Ч. Шеррингтоном (1906), обозначали все сенсорные приборы организма. Теперь же Проприоцепторы обычно рассматривают как одну из групп интероцепторов (см. Интероцепция). Проприоцепторы участвуют во всех актах, связанных с работой мышц, суставов и сухожилий (перемещение тела и его частей в пространстве, дыхание, процессы речеобразования и др.). С помощью П. организм получает информацию о характере перемещений тела, об особенностях стимулов (их величине, длительности, скорости, направлении и др.), действующих на опорно-двигательный аппарат. Благодаря деятельности П. возникают ощущения, названные И. М. Сеченовым «темным мышечным чувством».
Среди П. есть как обычные рецепторные образования (свободные нервные окончания, инкапсулированные рецепторы типа телец Руффини и Пачини и их разновидности), так и очень специфические сенсорные приборы, напр. сухожильные рецепторы Гольджи и мышечные веретена.
Свободные нервные окончания (см.) являются наиболее распространенным видом П. Инкапсулированные рецепторы чаще встречаются в мышечно-сухожильных соединениях и в тканях суставов (см. Механорецепторы). Сухожильные рецепторы располагаются в сухожилиях обычно на границе мышечной и сухожильной части мышцы, в опорных участках суставных капсул, в некапсульных и внутрикапсульных связках; имеют вытянутую форму. К ним подходят толстые афферентные мякотные волокна, а также более тонкие миелинизированные волоконца, заканчивающиеся, по мнению ряда ученых, свободными нервными окончаниями, оплетающими рецептор и, возможно, участвующими в возникновении ноцицептивной сигнализации.
Мышечные веретена имеют удлиненную структуру, расширяющуюся в средней части за счет капсулы, к-рая придает ей веретенообразную форму, что нашло отражение и в названии рецептора. У млекопитающих, в т. ч. у человека, эти рецепторы являются самыми сложными после специализированных органов чувств. Наибольшая плотность, оцениваемая по числу рецепторов на единицу веса мышцы (индекс Бокки), обнаружена в небольших мышцах рук, совершающих тонкие, высококоординированные движения. Как и во всех других рецепторах, в мышечных веретенах имеются вспомогательные структуры, представленные специализированными поперечнополосатыми мышечными волокнами (так наз. интрафузальные мышечные волокна), а также капсулой, содержимое к-рой по составу, по-видимому, отличается от обычной тканевой жидкости. В центре интрафузального волокна находится несократимый участок (область ядерной сумки). К нему с обеих сторон примыкают зоны измененной мышечной ткани (так наз. миотрубки), за к-рыми начинаются обычные мышечные структуры. Сенсорные нервные окончания мышечного веретена локализуются в области ядерной сумки и миотрубки. Выделяют два типа мышечных веретен: у мышечных веретен одного типа в центральной зоне интрафузальных волокон находится большое количество ядер (так наз. волокна с ядерной сумкой); у волокон другого типа (обычно более тонких и коротких) концентрация ядер в экваториальной зоне меньше, они располагаются в ряд, образуя цепочку (так наз. волокна с ядерной цепочкой). Этот тип волокон чаще встречается у животных, обладающих более тонкой координацией движений.
Интрафузальные волокна содержат три вида нервных окончаний. Два из них сенсорные (первичные, или аннулоспиральные, и вторичные, или гроздевидные), а один — моторный. Первичные окончания локализуются в экваториальной части волокон, вторичные же располагаются в области миотрубки. Моторные окончания рассеяны по остальным частям интрафузальных волокон. Первичные окончания у млекопитающих встречаются в волокнах обоих типов и образуются разветвлениями миелинезированных нервных волокон, которые являются наиболее крупными в организме (диаметр до 20 мкм) и относятся к волокнам группы Та. Вторичные окончания расположены обычно только в волокнах с ядерной цепочкой и образованы разветвлениями более тонких мякотных волокон группы П. Различия в диаметре нервных волокон, а также в скорости проведения импульсов по ним являются основными критериями различения сигналов, идущих от первичных и вторичных окончаний. Тонкая структура первичных и вторичных окончаний в целом одинакова. Эфферентные волокна (фузимоторные), направляющиеся в мышцу, представлены двумя основными группами: толстыми мякотными волокнами (альфа-группа), иннервирующими преимущественно экстрафузальные мышечные волокна, и тонкими волокнами (гамма-группа), иннервирующими интрафузальные волокна. Интрафузальные волокна иногда могут получать двигательную иннервацию и от волокон другого типа.
Проприоцепторы относятся к типу ареснитчатых механорецепторов. В основе механизма их возбуждения лежит деформация окончаний немиелинизированных разветвлений нервных волокон, следствием к-рой является изменение ионной проницаемости поверхностной мембраны механорецепторных терминалий. Между величиной статической фазы рецепторного потенциала и частотой импульсов существует прямо пропорциональная зависимость. Картина распределения импульсов при раздражении П. определяется рядом их свойств, таких как механические характеристики вспомогательных структур рецепторов, особенности рецепторного субстрата и связанной с ним системы генерации локальных электрических явлений (рецепторного, или генераторного, потенциала) и, наконец, особенности афферентных нервных волокон (прежде всего, свойства их начального сегмента, где под влиянием генераторного потенциала происходит возникновение потенциалов действия). У одних П. (напр., мышечных веретен сухожильных рецепторов, телец Руффини) хорошо выражен тонический компонент ответа, и эти П. являются медленно адаптирующимися рецепторами, воспринимающими статические компоненты раздражителей. У других П. (напр., телец Пачини) существует только динамическая составляющая реакции, и они являются быстро адаптирующимися рецепторами, хорошо воспринимающими такие воздействия, как быстрое движение и вибрация. Мышечные веретена располагаются в ткани параллельно экстрафузальным мышечным волокнам, тогда как сухожильные рецепторы — последовательно. Поэтому в момент мышечного сокращения нагрузка на мышечные веретена уменьшается, а на сухожильные рецепторы возрастает. При растяжении мышцы нагрузка на оба типа рецепторов возрастает, и все они возбуждаются. Эти особенности в расположении рецепторов приводят к тому, что мышечные веретена посылают в ц. н. с. информацию о длине мышцы, а сухожильные рецепторы — об ее натяжении (активном или пассивном). Соответственно, их называют датчиками длины и напряжения.
Эфферентная регуляция играет очень большую роль в работе Проприоцепторов. По эффективности обратной связи мышечные веретена занимают в ряду других тканевых рецепторов исключительное положение, что определяется их ролью в осуществлении быстропротекающих, строго координированных двигательных актов (см. Движения). Активизация эфферентных волокон приводит к сокращению интрафузальных мышечных волокон и, соответственно, к возбуждению самого рецептора. Все эфферентные гамма-волокна подразделяются на два типа: динамические и статические. Волокна первой группы обладают способностью возбуждать мышечное веретено в период динамического действия раздражения (растяжения мышцы). Волокна же второй группы действуют возбуждающим образом в период статической фазы растяжения. При постоянной длине мышцы раздражение волокон обоих типов вызывает возбуждение рецепторных окончаний. Статические фузимоторные волокна оказывают сходное действие на первичные и вторичные окончания. Влияние динамических волокон, по-видимому, ограничивается только первичными окончаниями. Гамма-волокна обычно обладают фоновой активностью, что приводит к флюктуации возбудимости нервных окончаний мышечного веретена. При этом первичные окончания, подверженные большему влиянию фузимоторной иннервации, демонстрируют большую вариабельность возбудимости.
Проприоцепторы являются важнейшим звеном в процессах осуществления самых разных двигательных актов (см. Движения), в поддержании мышечного тонуса (см.), сохранении позы (см.), равновесия тела (см.) и др. При перерезке афферентных путей от П. устраняется тоническое влияние импульсации от П. на мотонейроны спинного мозга, следствием чего является исчезновение мышечного тонуса. Кратковременное растяжение мышцы (напр., при ударе молоточком по ее сухожилию) вызывает всплеск активности П. и приводит к появлению миостатического рефлекторного ответа данной мышцы. Сухожильные рефлексы (см.) имеют кратчайшую (моносинаптическую) рефлекторную дугу и обусловлены активацией первичных нервных окончаний мышечных веретен. Эти рефлексы имеют большое значение в клин. практике при диагностике заболеваний спинного мозга (см.). К другой группе рефлексов, вызываемых активацией П., относятся статические реакции (см. Постуральные рефлексы). Они бывают локальными, сегментарными и генерализованными. Локальной реакцией является ответ мышцы на активацию собственных П. При длительном растяжении мышцы (десятки секунд) наблюдается тонический ответ. Так, растяжение мышцы на 1% повышает напряжение мышечных волокон на 2 кг. Сегментарными статическими реакциями являются ответы ц. н. с. и контралатеральных мышц на активацию П. других мышц этого сегмента спинного мозга. Типичным примером этих реакций служит экстензорный перекрестный рефлекс у децеребрированных животных. Сегментарные рефлексы играют важнейшую роль в процессах локомоции. Генерализованными статическими реакциями являются широко иррадиирующие ответы. При этом реакция мышц одних сегментов на раздражение П. других сегментов может модулироваться рефлекторными влияниями, обусловленными раздражением П. мышц шеи и рецепторов перепончатого лабиринта внутреннего уха. Совместно с П. в оценке движений, в формировании так наз. мышечного, или кинестетического, чувства участвуют и тактильные рецепторы кожи (см. Кожа, физиология). Все эти сенсорные структуры обычно рассматривают как периферическую часть сомато-кинестетического, или двигательного, анализатора (см. Двигательный анализатор). При различного рода поражениях периферических и центральных отделов нервной системы особенности нарушения мышечной и кожной чувствительности имеют важное диагностическое значение и, как правило, исследуются параллельно. При периферических поражениях зоны нарушения достаточно локальны; при спинномозговой патологии они могут носить сегментарный характер. Так, при односторонних поражениях спинного мозга наблюдается паралич Броун-Секара (см. Броун-Секара синдром), при к-ром на стороне поражения имеет место потеря мышечной чувствительности, а на противоположной стороне — кожной. Одним из наиболее постоянных признаков поражения задних корешков спинномозговых нервов на ранних стадиях спинной сухотки является значительное снижение мышечной чувствительности при незначительных изменениях тактильного и температурного чувства. Весьма значительные нарушения кинестетической и тактильной чувствительности возникают при поражении теменной области коры головного мозга.
Изучение Проприоцепторов осуществляется различными методами (с помощью световой и электронной микроскопии, методов электрофизиологии, биохимии и др.) При изучении П. у человека стали использовать метод отведения биопотенциалов от одиночных волокон с помощью микроэлектродов, вводимых в нервный ствол. Для анализа импульсных потоков широко применяют методы вычислительной техники.
См. также Рецепторы, Рецепция.
Библиография: Гранит Р. Основы регуляции движений, пер. с англ., М.. 1973; Ильинский О. Б. Физиология сенсорных систем, ч. 3 — Физиология механорецепторов, Л., 1975; Шеррингтон Ч. Интегративная деятельность нервной системы, пер. с англ., Л., 1969; Handbook of sensory physiology, ed. by С. C. Hunt, v. 3/2, B., 1974; Matthews P. В. C. Mammalian muscle receptors and their central actions, Baltimore, 1972.
О. Б. Ильинский.
- Haeser, «Handbuch der Gesch. d. Medicin».
- Frédault, «Histoire de la médecine» (П., 1970).
- Sprengel, «Pragmatische Geschichte der Heilkunde».
- https://scorcher.ru/axiomatics/axiom_show.php?id=314.
- https://allasamsonova.ru/receptory-skeletnoj-myshcy-myshechnye-veretena/.
- https://xn--90aw5c.xn--c1avg/index.php/%D0%9F%D0%A0%D0%9E%D0%9F%D0%A0%D0%98%D0%9E%D0%A6%D0%95%D0%9F%D0%A2%D0%9E%D0%A0%D0%AB.
- Haeser, «Handbuch der Gesch. d. Medicin».

")







