
Можно выделить два основных вида специфических рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи). В этой статье рассмотрено строение и функции одного вида рецепторов скелетной мышцы — мышечных веретен. Описаны строение и функции интрафузальных мышечных волокон, входящих в состав мышечных веретен.
РЕЦЕПТОРЫ СКЕЛЕТНОЙ МЫШЦЫ (МЫШЕЧНЫЕ ВЕРЕТЕНА)
Познакомившись с строением нервов, можно обсудить вопрос о рецепторах скелетной мышцы. Можно выделить два основных вида рецепторов, расположенных в скелетной мышце: мышечные веретена и рецепторы Гольджи (сухожильные рецепторы Гольджи).
Мышечные веретена были описаны в середине XIX века немецким физиологом Вильгельмом Кюне (Wilhelm Kühne, 1863). Мышечное веретено представляет собой вытянутую структуру, расширенную посередине, что делает его похожим на веретено, которое в давние времена использовали, когда пряли. Английский физиолог, лауреат Нобелевской премии Чарльз Скотт Шеррингтон одним из первых указал, что мышечные веретена являются чувствительными рецепторами мышцы.
Мышечные веретена расположены внутри мышцы параллельно мышечным волокнам и прикрепляются к эндомизию, окружающему мышечные волокна или к сухожилию. Длина мышечного веретена достигает 10 мм. Ширина мышечного веретена в наиболее широкой его части — капсуле колеблется от 80 до 250 мкм (рис.1)
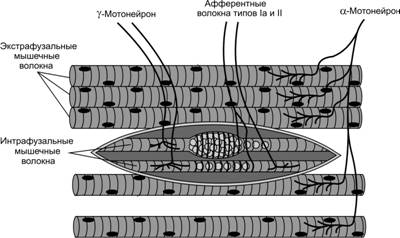
Рис.1. Строение мышечного веретена
Более подробно строение и функции мышц описаны в моих книгах
Количество мышечных веретен в мышце колеблется от нескольких десятков до нескольких сотен. Так, в двуглавой мышце плеча имеется 320 веретен, в большой грудной — 450, а в трехглавой мышце плеча — 520. Плотность веретен (то есть количество веретен приходящихся на 1 г массы мышцы) сильно варьирует. Она наибольшая в тыльных мышцах шеи и наименьшая в мышцах конечностей. Так, например в верхней косой мышце головы плотность веретен составляет 42,7 на 1 г массы мышцы, а в двуглавой мышце плеча — 2,0 на 1 г массы мышцы.
Установлено, что мышечные веретена возбуждаются при растягивании мышцы. Растяжение мышцы возникает, когда, например, занимающиеся выполняют упражнения на растяжение или упражнения в уступающем (эксцентрическом режиме). Также растяжение мышц возникает при локомоциях человека — ходьбе, беге, прыжках или езде на велосипеде.
Информация, поступающая от рецепторов мышечных веретен, сообщает нервной системе о том, какова в настоящий момент времени фактическая длина мышцы (ее еще называют текущей длиной — то есть длиной в текущий момент времени) и скорость растяжения мышцы.
Внутри мышечного веретена располагаются тонкие мышечные волокна, которые называются интрафузальными. Их диаметр в 2-3 раза меньше обычных мышечных волокон, называемых экстрафузальными. Интрафузальные мышечные волокна очень маленькие, однако они также могут сокращаться и расслабляться. Интрафузальных мышечных волокон в мышечном веретене немного: от 4 до 14. Следует отметить, что интрафузальные мышечные волокна в веретене имеют собственную иннервацию, которая позволяет регулировать чувствительность мышечного веретена. К интрафузальным мышечным волокнам подходят двигательные мотонейроны (γ-мотонейроны) и от них отходят чувствительные волокна, несущие информацию в ЦНС о длине и скорости растяжения мышцы. За открытия в этой области шведскому физиологу Рагнару Граниту была присуждена Нобелевская премия.
Благодаря информации, поступающей от мышечных веретен, предотвращается (правда не всегда) повреждение мышцы. Эта информация служит основой для функционирования рефлекса на растяжение.
Литература
- Гранит Р. Основы регуляции движений. — М.: Мир, 1973.- 278 с. Ил.
- Мак-Комас, А.Дж. Скелетные мышцы. — Киев: Олимпийская литература, 2001.- 407 с.
- Самсонова А.В. Моторные и сенсорные компоненты биомеханической структуры физических упражнений: автореф. дис… докт. пед. наук.- СПб, 1998.- 48 с.
- Самсонова, А.В. Биомеханика мышц [Текст]: учебно-методическое пособие /А.В. Самсонова Е.Н. Комиссарова /Под ред. А.В. Самсоновой /Санкт-Петербургский гос. Ун-т физической культуры им. П.Ф. Лесгафта.- СПб,: [б.н.], 2008.- 127 с.
- Самсонова, А.В. Гипертрофия скелетных мышц человека. — СПб: Кинетика, 2018.- 159 с.
С уважением, А.В.Самсонова
Похожие записи:
Томас ДеЛорме — ученый, разработавший метод прогрессивно возрастающего сопротивления
Описаны достижения врача-реабилитолога Томаса ДеЛорме, который разработал специальные силовые тренажеры для своих пациентов; использовал большие отягощения для борьбы…
Мышечные боли или почему болят мышцы после тренировок? (запаздывающие болезненные ощущения)
Описаны виды и причины болей в мышцах, возникающих через день после тренировки. Показано, что причиной мышечных болей…
Мышечные боли или почему болят мышцы во время и после тренировок? (острые болезненные ощущения)
Описаны виды и причины болей в мышцах после тренировки. Основное внимание уделено мышечным болям, возникающим во время…
От чего зависит сила мышц? (физиологические факторы)
Описаны физиологические факторы, определяющие силу скелетных мышц человека: частота импульсации ДЕ, количество активных ДЕ, синхронизация…
Классификация соматотипов детей по Штефко-Островскому
Описана классификация типов телосложения детей по Штефко-Островскому: астеноидного, торакального, мышечного и дигестивного. Кратко описан жизненный путь В.Г. Штефко.
Типы телосложения (соматотипы) по Хит-Картеру
Описаны типы телосложения (соматотипы) по классификации Б.Х. Хит и Д.Э.Л. Картера. Показаны преимущества разработанной классификации по сравнению…
Мышечные рецепторы.Регуляция работы мышечного веретена.Сухожильные органы Гольджи.
• Мышечные веретена — рецепторы растяжения мышц, бывают двух типов: o с ядерной сумкой o с ядерной цепочкой • Сухожильный орган Гольджи — рецепторы сокращения мышц. При сокращении мышцы сухожилие растягивается и его волокна пережимают рецепторное окончание, активируя его. Нервно-мышечное веретено — сложный рецептор, который включает видоизмененные мышечные клетки, афферентные и эфферентные нервные отростки и контролирует как скорость, так и степень сокращения и растяжение скелетных мышц. Строение мышечных веретен В каждой поперечнополосатой мышце содержатся мышечные веретена. Мышечные веретена, в соответствии с названием, имеют форму веретена длиной несколько миллиметров и диаметром несколько десятых долей миллиметра. Веретена расположены в толще мышцы параллельно обычным мышечным волокнам. Мышечное веретено имеет соединительнотканную капсулу. Капсула обеспечивает механическую защиту элементов веретена, расположенных в полости капсулы, регулирует химическую жидкую среду этих элементов и этим обеспечивает их взаимодействие. В полости капсулы мышечного веретена расположено несколько особых мышечных волокон, способных к сокращению, но отличающихся от обычных мышечных волокон мышцы как по строению, так и по функции. Эти мышечные волокна, расположенные внутри капсулы, назвали интрафузальными мышечными волокнами (лат.: intra — внутри; fusus — веретено); обычные мышечные волокна называются экстрафузальными мышечными волокнами (лат.: — вне, снаружи; fusus — веретено). Интрафузальные мышечные волокна тоньше и короче экстрафузальных мышечных волокон. Выделяют два главных типа интрафузалъных мышечных волокон. Один тип интрафузального мышечного волокона — волокно с ядерной сумкой (bursa nuclearis)Это волокно имеет в средней части около сотни компактно собранных клеточных ядер. Утолщенная средняя часть такого интрафузального волокна представляет собой сумку с ядрами. Именно потому эти волокна назвали сумчато-ядерными. Другой тип волокна, волокно с ядерной цепочкой (vinculun nucleare) , имеет ядра распределенные в виде цепочки по длинной оси интрафузального волокна. Именно потому эти волокна называют цепочно-ядерными интрафузальными мышечными волокнами. Цепочечно-ядерные волокна вдвое тоньше и почти вдвое короче, чем сумчато-ядерные волокна. 20!!!!!!!!!!!!!!!!!!!!!!!!!!!!!! Иннервация мышечных веретен Афферентная иннервация В полость каждого мышечного веретена на уровне ядерной зоны проникают нервные волокна и кровеносные сосуды. Среди нервных волокон одно — толстое миелинизированное нервное волокно. Внутри мышечного веретена одиночное крупное волокно ветвится и посылает терминали дендритов к каждому интрафузальному мышечному волокну любых разновидностей. Конечные нервные ветви обвиваются вокруг средних частей интрафузальных волокон, образуя так называемое аннулоспиральное окончание (лат.: anulus — колечко, завиток; anulo- в форме кольца). Афферентные нервные волокна, образующие аннулоспиральные окончания, принадлежат к типу Iа. Их называют первичными афферентными волокнами мышечных веретен. В соответствии с этим аннулоспиральные нервные окончания носят название первичных чувствительных (сенсорных) окончаний. Полагают, что каждое мышечное веретено иннервируется только одним афферентным нервным волокном типа Iа. Большинство мышечных веретен иннервируются также одним или несколькими афферентными волокнами типа II. Они могут иннервировать все разновидности интрафузальных мышечных волокон, но главным образом иннервируют цепочечноядерные интрафузальные мышечные волокна. Терминали афферентных волокон типа II располагаются в краевых участках интрафузальных мышечных волокон к периферии от аннулоспиральных окончаний. Такие структуры называют вторичными сенсорными окончаниями. Они могут либо обвивать цепочечно-ядерное интрафузальное волокно в виде нерегулярной спиральной пружины, или образовывать множественные разветвления в сумчато-ядерном волокне, называемые «гроздевидными окончаниями». В отличие от волокон Iа, волокна группы II часто иннервируют два или несколько мышечных веретен. Эфферентная иннервация Как экстрафузальные, так и интрафузальные мышечные волокна иннервируются эфферентными нервными волокнами, аксонами мотонейронов спинного мозга. Это разные мотонейроны. Экстрафузальные волокна иннервируются α-мотонейронами, а интрафузальные — γ-мотонейронами. Тела γ-мотонейронов, посылающих по своим аксонам (фузимоторные нервные волокна) управляющие сигналы к мышечным веретенам, значительно меньше по размерам, чем тела α-мотонейронов, управляющих экстрафузальными мышечными волокнами. Фузимоторные нервные волокна значительно тоньше эфферентных волокон, управляющих сокращениями мышц. Фузимоторные нервные волокна называют γ-(эфферентными) нервными волокнами. В пределах мышцы γ-волокна разветвляются и иннервируют несколько мышечных веретен. Внутри каждого веретена γ-волокна иннервируют несколько интрафузальных мышечных волокон. γ-волокна образуют несколько типов окончаний на периферических (полярных) участках интрафузальных мышечных волокон. Эти окончания называют γ-концевыми пластинками, если они локализованы на сумчато-ядерных интрафузальных мышечных волокнах, а также γ-кустовидными нервными окончаниями, если они локализованы на цепочечноядерных интрафузальных мышечных волокнах. γ-концевые пластинки подобны обычным концевыми пластинкам, расположенным на экстрафузальных мышечных волокнах). γ-кустовидные окончания представляют собой длинные тонкие структуры, разветвленные в виде диффузной сети. Каждое γ-волокно образует только один тип терминалей: либо только кустовидные окончания, либо только концевые пластинки. Приходящие по эфферентным нервным окончаниям импульсы вызывают сокращение мышечных волокон, что ведёт к деформации афферентных окончаний и приводит к возникновению нервных импульсов, частота которых пропорциональна скорость изменения длины мышечных волокон и величине этого изменения. Сухожильный орган Гольджи (нервно-сухожильное веретено) — рецепторный орган, располагающийся в местах соединения мышечных волокон с коллагеновыми пучками сухожилий. Весь орган окружён тонкой соединительнотканной капсулой. Нервное волокно, подходящее к веретену утрачивает миелиновый слой и, распадаясь на терминальные ветви, проходит между пучками коллагеновых волокон сухожилия. Сокращаясь, мышца растягивает сухожилие. Это вызывает возбуждение нернвых окончаний чувствительных нейронов спинного мозга. Они возбуждают тормозные нейроны спинного мозга, которые тормозят соответствующие двигательные нейроны, что предотвращает перерастяжения мышцы. Известно, что порог возбуждения рецептеров сухожильного органа Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен. Это позволяет мышце сокращаться без всяких помех до определенного предела 20. Моно- и полисинаптические рефлексы спинного мозга. Миотатический и обратный миотатический рефлекс. 21. моно и полисинаптические рефлексы спинного мозга. Миотатический и обратный миотатический рефлекс. Спинной мозг — часть центральной нервной системы, расположенная в позвоночном канале. Рефлекс (от лат reflexus — отраженный) — стереотипная реакция организма на определенное воздействие, проходящая с участием нервной системы. Рефлексы существуют у многоклеточных живых организмов, обладающих нервной системой. По степени сложности нейронной организации рефлекторные дуг различают моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный), и полисинаптические, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений (например, флексорный). Существуют и более сложные рефлекторные дуги, включающие один или неск. вставочных нейронов между афферентной и эфферентной частями рефлекса. Примером простейшего полисинаптического (более чем с одним синапсом) рефлекса может служить сухожильный рефлекс. Сенсорный концевой орган — тельца Гольджи — находится в сухожилиях. Увеличение нагрузки на сухожилие, вызываемое обычно сокращением прикрепленной к нему мышцы, и есть возбуждающий раздражитель, к-рый приводит к растяжению телец Гольджи и возникновению в них импульсной активности, распростр. по соотв. афферентному волокну. Идущий от сухожильного сенсорного концевого органа афферент заканчивается на вставочном нейроне в спинном мозге. Этот вставочный нейрон оказывает тормозящее действие на α-мотонейрон, понижая активность в его эфферентном аксоне. Поскольку этот аксон возвращается к мышце, прикрепленной к растянутому сухожилию, мышца расслабляется и нагрузка на сухожилие снижается. Рефлекс растяжения мышцы и сухожильный рефлекс работают во взаимодействии, обеспечивая базисный механизм быстрого регулирования степени сокращения мышцы. Эти Р. полезны для быстрых приспособлений к изменению положения ноги, когда чел. приходится идти по неровной почве. Разумеется, в локомоции участвуют и др. полисинаптические спинальные Р. Эти Р. включают гораздо больше вставочных нейронов в структуру рефлекторной дуги. Неврологическую основу этих сложных Р. образуют дивергентные (от одного нейрона к неск.) и конвергентные (от неск. нейронов к одному) связи вставочных нейронов. Моносинаптические и полисинаптические спинномозговые Р. образуют базовый механизм поддержания и приспособления позы. Моторные системы головного мозга влияют на спинномозговые Р. через входные цепи, идущие к вставочным нейронам и γ-мотонейронам. Т. о., изменения спинномозговых Р. могут указывать на патологию в моторных системах головного мозга. Примером этого может служить гиперрефлексия, связанная с травмой латеральных спинномозговых двигательных путей или с повреждением моторных областей лобной доли. Простейшим рефлексом яв-ся миотатический рефлекс, или рефлекс растяжения мышцы. Этот рефлекс можно вызвать у любой скелетной мышцы, хотя самый известный его пример — коленный рефлекс. Анат. основу миотатического рефлекса составляет моносинаптическая (с одним синапсом) рефлекторная дуга. Она включает сенсорный концевой орган, сенсорное нервное волокно с его клеточным телом в ганглии заднего корешка, α-мотонейрон, на к-ром сенсорный аксон образует синапс, и аксон этого α-мотонейрона, возвращающийся к мышце, от к-рой приходит сенсорное волокно. Сенсорным концевым органом в рефлексе растяжения мышцы служит мышечное веретено. Мышечное веретено имеет мышечные окончания, наз. интрафузальными волокнами, и центральную, немышечную область, связанную с окончанием афферентного нерва. Интрафузальные волокна иннервируются γ-мотонейронами передних корешков спинного мозга. Высшие центры головного мозга могут оказывать влияние на рефлекс растяжения мышцы посредством модуляции активности γ-мотонейронов. Этот рефлекс вызывается растяжением мышцы, что приводит к увеличению длины мышечного веретена и, вследствие этого, к повышению частоты генерирования потенциала действия в сенсорном (афферентном) нервном волокне. Повышенная активность в афферентном волокне усиливает разряд целевого γ-мотонейрона, что вызывает сокращение экстрафузальных волокон мышцы, от к-рой приходит афферентный сигнал. Когда экстрафузальные волокна сокращаются, мышца укорачивается и активность в афферентных волокнах понижается 21. |
Структура и функции проприорецепторов
В активной части опорно двигательного аппарата имеются проприорецепторы. Существует два типа проприорецепторов — нервно-мышечные веретёна, которые реагируют на изменение длины мышцы, а также сухожильные органы Гольджи, реагирующие на изменение напряжения, то есть на силу сокращения мышечных волокон (Рисунок 1).

Рисунок 1. Проприорецепторы: нервно-мышечное веретено и сухожильный орган Гольджи
Нервно-мышечные веретёна. Нервно-мышечные веретена расположены в толще скелетных мышц между мышечными волокнами. Они представляют собой веретеновидные структуры, покрытые соединительнотканной капсулой. Длина веретена составляет 4 — 7 (до 10) мм, диаметр — 100 мкм. Под капсулой внутри веретена находятся видоизменённые мышечные волокна, которые называются интрафузальными. Эти волокна имеют ряд отличий от обычных, экстрафузальных, мышечных волокон. Во-первых, интрафузальное волокно имеет меньший диаметр (15 — 30 мкм) по сравнению с экстрафузальным (50 — 100 мкм). Во-вторых, в центральной («экваториальной») части интрафузального волокна нет сократительных элементов, поэтому к сокращению способны только периферические части. Поскольку концы интрафузального волокна фиксированы к соединительнотканной капсуле, покрывающей веретено, при укорочении периферических отделов волокна его центральная часть растягивается. Интрафузальные волокна расположены параллельно эктсрафузальным. По строению интрафузальные волокна делятся на два типа — с ядерной сумкой и с ядерной цепочкой (Рисунок 2). В свою очередь, волокна с ядерной сумкой по функции по особенностям иннервации подразделяются на статические и динамические. Для волокон с ядерной сумкой характерно наличие расширенной центральной части, в которой ядра собраны в скопления. Волокна с ядерной цепочкой примерно вдвое тоньше волокон с ядерной сумкой, ядра в их центральной части расположены в ряд. В каждом веретене содержится 1 — 3 волокна с ядерной сумкой и 5 — 7 волокон с ядерной цепочкой.

Рисунок 2. Нервно-мышечное веретено
Интрафузальные волокна получают как афферентную, так и эфферентную иннервацию. Выделяют два типа афферентных окончаний — первичные и вторичные. Первичные окончания называют аннулоспиральными, они представляют собой спираль, окружающую центральную часть всех интрафузальных волокон. От первичных окончаний отходят волокна типа Ia, которые соответствуют типу Aα по Гассеру-Эрлангеру. Это самые толстые волокна (~17 мкм) с самой высокой скоростью проведения возбуждения (70 — 12 м/с). Каждое нервно-мышечное веретено иннервируется только одним волокном Ia. Другими словами одно афферентное нервное волокно Ia получает импульсацию со всех первичных окончаний в иннервируемом им веретене. Вторичные окончания имеют форму спирали, но могут также образовывать разветвлённую структуру. Они оплетают центральную часть интрафузальных волокон с ядерной цепочкой, а также статических волокон с ядерной сумкой, но их не бывает в динамических волокнах с ядерной сумкой. От вторичных окончаний отходят волокна типа II, соответствующие типу Аβ по Гассеру-Эрлангеру; диаметр этих волокон составляет ~8 мкм, они проводят возбуждение со скоростью 40 — 70 м/с (Рисунок 3).

Рисунок 3. Схема нервно-мышечного веретена
Как первичные, так и вторичные нервные окончания чувствительны к изменению длины интрафузальных волокон, однако характер ответной реакции различается. При медленном растяжении центральной части интрафузальных волокон импульсация появляется как в первичных, так и во вторичных рецепторах. Частота импульсов пропорциональная степени растяжения волокна. Такой ответ афферентных окончаний называют статическим. Однако при быстром внезапном растяжении волокна в первичных (но не вторичных) окончаниях отмечается мощный всплеск импульсации — динамический ответ. Следовательно, волокна Ia передают информацию как об абсолютной длине интрафузальных волокон (то есть о степени растяжения), так о скорости её изменения, тогда как волокна II — только об абсолютной длине. Следует понимать, что растяжение центральной части интрафузальных волокон может быть связано как с удлинением всей мышцы (интрафузальные волокна расположены параллельно экстарфузальным), так и при сокращении периферических отделов волокна (Рисунок 3).

Рисунок 4. Статический и динамический ответы рецепторов нервно-мышечных веретён
Эфферентная иннервация интрафузальных волокон представлена нервными волокнами типа Aγ, которые являются аксонами γ-мотонейронов спинного мозга. Диаметр этих волокон составляет ~5 мкм, скорость проведения возбуждения в них — 15 — 40 м/с. Окончания Aγ волокон подходят к периферическим отделам интрафузальных волокон, при возбуждении γ-мотонейронов концевые участки интрафузальных волокон сокращаются, в результате чего их центральные отделы растягиваются. Выделяют γ-динамические и γстатические волокна, γ-динамические волокна иннервируют динамичекие волокна с ядерной сумкой, их активация повышает динамическую чувствительность окончаний Iа типа.
γ-статические волокна иннервируют волокна с ядерной цепочкой и статические волокна с ядерной сумкой, их возбуждение повышает уровень активности как первичных, так и вторичных окончаний (см. ниже).
Миотатический рефлекс (рефлекс растяжения)
В ответ на увеличение длины мышцы рецепторы нервно-мышечных веретён возбуждаются, и импульсация по волокнам Ia (и при быстром, и при медленном растяжении) или II (при медленном растяжении) поступает в спинной мозг. Войдя в серое вещество спинного мозга, волокно афферентного нейрона разветвляется и образует возбуждающие синапсы практически со всеми α-моторными нейронами, иннервирующими гомонимную (то есть эту же) мышцу. Возбуждение α-моторных нейронов вызывает сокращение экстрафузальных волокон, и мышца укорачивается. Следует отметить, что коллатерали афферентных нейронов переключаются также на вставочные тормозные нейроны, которые угнетают α-мотонейроны мышц антагонистов. Таким образом, растяжение мышцы вызывает её рефлекторное укорочение, при этом мышцы антагонисты расслабляются. Описанный рефлекс с рецепторов нервно-мышечных веретён называется миотатическим, или рефлексом на растяжение (Рисунок 4). Миотатический рефлекс является моносинаптическим, поэтому имеет очень небольшой латентный период (~ 0,9 мс).

Рисунок 5. Миотатический рефлекс с нервно-мышечных веретён
Различают фазический (динамический) и тонический миотатический рефлекс. Фазический рефлекс запускается с первичных нервных окончаний при быстром растяжении мышцы, например, при ударе по сухожилию неврологическим молоточком или при появлении внезапного препятствия совершаемому движению. Тонический рефлекс запускается с вторичных нервных окончаний при медленном (или постоянном) растяжении мышечных волокон; он имеет значение для поддержания вертикальной позы. Когда тело отклоняется от правильного положения, мышцы, участвующие в поддержании позы, растягиваются. В результате возбуждаются как первичные, так и вторичные окончания нервно-мышечных веретён, запускается миотатический рефлекс, и мышцы сокращаются; за счёт их сокращения тело возвращается в правильное положение. Когда у длительно стоящего человека (например, солдата в стойке «смирно») из-за утомления сгибаются ноги в коленных суставах, проприорецепторы четырёхглавых мышц реагируют на растяжение, и ноги снова выпрямляются. Постоянное стимулирующее влияние на α-мотонейроны со стороны афферентных волокон Ia и II типов вносит вклад в поддержание мышечного тонуса.
Функции γ-моторных нейронов. Понятие о γ-петле.
Как уже упоминалось, интрафузальные волокна получают эфферентную иннервацию со стороны γ-моторных нейронов. При возбуждении γ-мотонейронов происходит сокращение периферических (концевых) отделов интрафузальных волокон, в результате их центральная часть растягивается. При этом первичные и вторичные нервные окончания активируются, импульсация от них поступает в спинной мозг, и возбуждаются α-моторные нейроны. Другими словами, запускается миотатический рефлекс. Такой механизм (активация рефлекса растяжения при возбуждении γ-мотонейронов) называется γ-петлёй. Наличие γ-петли обеспечивает сохранение чувствительности рецепторов нервномышечных веретён при разной длине мышцы. Во время сокращения мышцы её длина уменьшается, соответственно должны укорачиваться и интрафузальные волокна. В результате импульсация с рецепторов нервно-мышечных веретён должна была бы прекратиться, и рецепторы потеряли бы способность реагировать на изменение длины мышцы. Напротив, при расслаблении мышцы её длина увеличивается, и интрафузальные волокна могут быть чрезмерно растянуты. В таких условиях рецепторы будут генерировать импульсы с максимальной частотой и, следовательно, также потеряют способность реагировать на изменение длины. Однако в реальных условиях имеет место α-γ-коактивация, то есть α- и γмотонейроны возбуждаются одновременно. В результате сокращаются как экстра-, так и интрафузальные волокна. Сокращение экстрафузальных волокон приводит к укорочению мышцы, а сокращение периферических отделов интрафузальных волокон — к растяжению их центральной части. Следовательно, длина центральной части не меняется, поскольку её укорочение вследствие сокращения экстрафузальных волокон компенсируется растяжением из-за сокращения периферических отделов интрафузальных волокон. И наоборот — расслабление всей мышцы сопровождается расслаблением и интрафузальных волокон, при этом растяжение центральной части уменьшается. Таким образом, за счёт α-γ-коактивации рецепторы нервно-мышечных веретён сохраняют чувствительность (и, следовательно, передают информацию о длине мышцы в ЦНС) как при сокращении мышцы, так и при её расслаблении (Рисунок 6). На γ-мотонейронах заканчиваются нисходящие пути из различных отделов головного мозга. Меняя активность γ-мотонейронов, мозговые центры влияют на чувствительность рецепторов нервно-мышечных веретён и, следовательно, на порог рефлекса растяжения. Сухожильные органы Гольджи. Обратный миотатический рефлекс Кроме нервно-мышечных веретён в активной части опорно-двигательного аппарата имеется ещё один тип проприрецепторов — сухожильные органы Гольджи. Сухожильные органы представляют собой сеть коллагеновых нитей, окружённую тонкой соединительнотканной капсулой. Диаметр сухожильного органа Гольджи составляет 100 мкм, длина — 1 мм. Коллагеновые нити внутри органа оплетённых нервными окончаниями, от которых отходят нервные волокна типа Ib. эти нервные волокна, как и Ia, соответствуют типу Aα по Гассеру-Эрлангеру. Сухожильные органы Гольджи расположены в месте прикрепления мышечных волокон к сухожильным, при этом каждый сухожильный орган связан с 3 — 25 (в среднем — с 10 — 15) мышечными волокнами (Рисунок 7). Таким образом, в отличии от нервно-мышечных веретён, которые ориентированы параллельно мышечным
волокнам, сухожильные органы по отношению к мышечным волокнам расположены последовательно.

Рисунок 6. Функция γ-петли
Следовательно, сухожильные органы реагируют не на изменение длины мышцы, а на развиваемое ей усилие, другими словами, на степень её напряжения. Афферентные нервные волокна от сухожильных органов Гольджи следуют в спинной мозг, где, в отличие от чувствительных волокон с нервно-мышечных веретён, переключаются на вставочные тормозные нейроны. Вставочные нейроны, в свою очередь, образуют синаптические контакты с α-мотонейронами гомонимной мышцы. Таким образом, активация сухожильных органов приводит к расслаблению мышцы, к которой крепится данное сухожилие. Описанный рефлекс называется обратным рефлексом растяжения (обратным миотатическим рефлексом) (Рисунок 8). Этот рефлекс, по-видимому, имеет две функции. Во-первых, он предупреждает чрезмерное напряжение скелетных мышц, которое может привести к тяжёлым травмам (разрыву мышцы, отрыву сухожилия от кости или от мышцы). Во-вторых, обратный миотатический рефлекс обеспечивает равномерность силы сокращения разных мышечных волокон в пределах одной мышцы. Те волокна, которые напряжены слишком сильно, расслабляются за счёт усиления тормозных влияний со стороны спинного мозга. Напряжение волокон, которые сокращаются недостаточно сильно, напротив, усиливается за счёт уменьшения тормозного воздействия.

Рисунок 7. Сухожильный орган Гольджи
Проприорецепторы посылают в головной мозг информацию о состоянии скелетно-мышечной системы Импульсация с проприорецепторов поступает не только в спинной мозг, но и по коллатералям афферентных волокон в различные отделы головного мозга. В частности, импульсы с нервно-мышечных веретён и сухожильных органов следуют по спинномозжечковым трактам в мозжечок, а также по системе задних столбов (тонкий и клиновидный пучок) в продолговатый мозг, затем — в таламус, а оттуда — в кору больших полушарий. Таким образом, мозжечок и кора большого мозга постоянно получают информацию о состоянии скелетно-мышечной системы, а именно о длине мышц и о степени их напряжения. Эта информация является необходимой как для регуляции произвольных движений и позы, так и для осознания положения частей тела в пространстве и друг относительно друга.

Рисунок 8. Обратный миотатический рефлекс с сухожильных органов Гольджи
- М.П. Киселева, З.С. Смирнова, Л.М. Борисова и др. Поиск новых противоопухолевых соединений среди производных N-гликозидов индоло[2,3-а] карбазолов // Российский онкологический журнал. 2015. № 1. С. 33-37.
- Frédault, «Histoire de la médecine» (П., 1970).
- Frédault, «Histoire de la médecine» (П., 1970).
- https://allasamsonova.ru/receptory-skeletnoj-myshcy-myshechnye-veretena/.
- https://cyberpedia.su/17x41cd.html.
- https://manumed.pro/%D1%81%D1%82%D1%80%D1%83%D0%BA%D1%82%D1%83%D1%80%D0%B0-%D0%B8-%D1%84%D1%83%D0%BD%D0%BA%D1%86%D0%B8%D0%B8-%D0%BF%D1%80%D0%BE%D0%BF%D1%80%D0%B8%D0%BE%D1%80%D0%B5%D1%86%D0%B5%D0%BF%D1%82%D0%BE%D1%80/.
- Patil H., Tiwari R. V., Repka M. A. Recent advancements in mucoadhesive floating drug delivery systems: A mini-review. Journal of Drug Delivery Science and Technology. 2016; 31: 65–71.DOI: 10.1016/j.jddst.2015.12.002.
- ОФС.1.2.1.2.0003.15 Тонкослойная хроматография // Государственная фармакопея, XIII изд.
- Bangun H., Aulia F., Arianto A., Nainggolan M. Preparation of mucoadhesive gastroretentive drug delivery system of alginate beads containing turmeric extract and anti-gastric ulcer activity. Asian Journal of Pharmaceutical and Clinical Research. 2019; 12(1):316–320. DOI: 10.22159/ajpcr.2019.v12i1.29715.
- Puccinotti, «Storia della medicina» (Ливорно, 1954—1959).

")







